

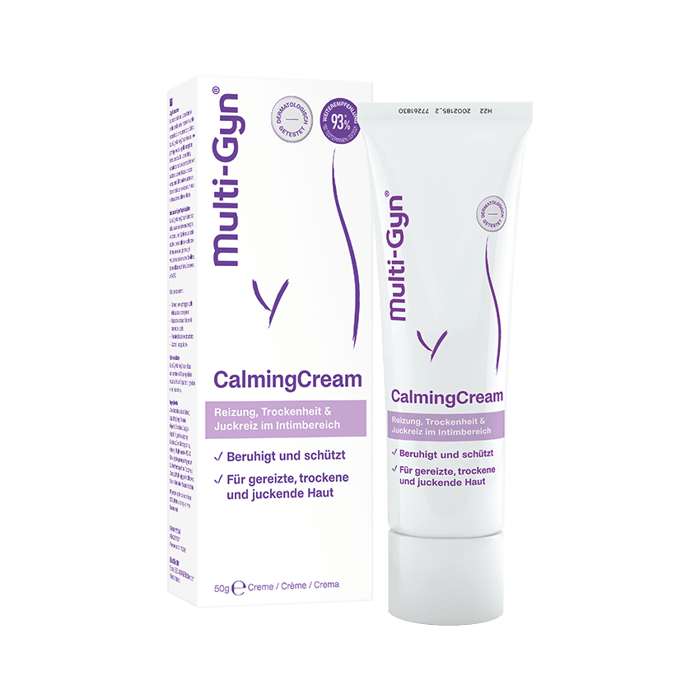



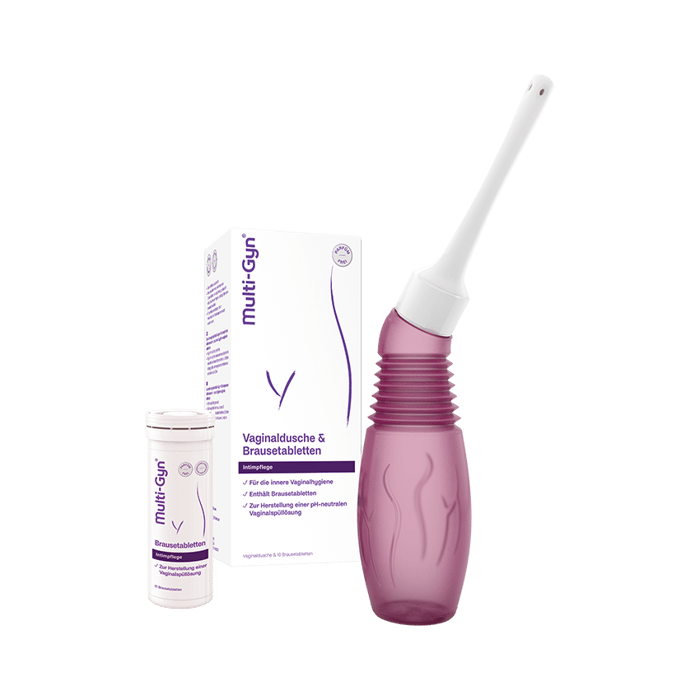


Dieses Problem betrifft uns alle mal
Wir alle genießen unser Leben, aber jede Dritte von uns leidet an einer bakteriellen Vaginose, ohne zu wissen, was das ist oder wie man sie behandelt. Du bist Dir noch unsicher, was Deine Beschwerden verursacht? Wir können helfen.